Двенадцатиперстная кишка (duodenum) — передний отдел средней кишки. Она начинается около пилорической железы, идет вниз до середины полости тела и, загибаясь вверх, дает восходящую петлю, которая поднимается до уровня нижнего края правой доли печени или даже до пилорической железы, снова поворачивает налево и переходит в расширенную часть — спиральную кишку.
Вскрывают продольным разрезом стенку кишечника, рассматривают строение внутренних стенок. Они покрыты слизистой оболочкой, имеющей своеобразную структуру, из-за наличия углубленных ячеек. Отверстие пилорической железы лежит рядом со сфинктером пилорического желудка. Тут же впадает желчный проток (ductus choledochus) желчного пузыря.
Селезенка (lien,) у осетровых рыб — большой железистый орган V-образной формы, огибающий справа и слева петлю двенадцатиперстной кишки и подстилающий ее, что можно видеть, приподняв кишку. Начинается селезенка отдельной долей, примыкающей к пилорической железе, и далее переходит в удлиненную долю, которая образует внизу выступ в форме клина. Затем она поднимается по восходящей петле до края правой доли печени, где уже с правой стороны, под нижней долей печени, образует отдельную треугольную долю, называемую добавочной долей селезенки. Эта добавочная доля прилегает непосредственно к печени и поджелудочной железе, так что границу между ними можно определить лишь по цвету желез. У осетра добавочная доля достигает значительных размеров. Если отвернуть петлю двенадцатиперстной кишки, то можно увидеть, что селезенка, располагающаяся под ней, имеет Z-образную форму и подстилает кишку справа и слева.
Поджелудочная железа (pancreas) у осетровых рыб вместе с Жаровыми скоплениями располагается вдоль сосудов, питающих двенадцатиперстную кишку, и находится в тесном контакте с лопастями правой доли печени и дополнительной долей селезенки. Поэтому нередко печень осетровых рыб называют hepatopancreas. Однако (особенно у крупных осетров) можно различить отдельные лопасти поджелудочной железы. Они располагаются на месте перехода пилорического отдела желудка в двенадцатиперстную кишку и тянутся двумя продольными лопастями: одна лежит вдоль наружной стенки вместе с подкишечной веной (v. intestinalis), другая — по внутреннему краю вдоль кишечной артерии (a. mesenterica). Обе лопасти подходят к центральной доле печени.
Спиральная кишка (colon) — задний отдел средней кишки, нисходящий отдел кишечника — является непосредственным продолжением вниз двенадцатиперстной кишки. Этот отдел кишечника имеет толстые мускулистые стенки и даже с наружной стороны его видны внутрилежащие складки спирального клапана. Спиральный клапан образуется скрученностью кишечной трубки и созданием внутри нее закрученной высокой складки слизистой оболочки, свисающей в полость, что можно видеть на поперечном разрезе, Всего имеется 7-8 таких витков. На вскрытой кишечной трубке создается впечатление, что она разделена на 7-8 отдельных камер, сообщающихся друг с другом. Помимо спирального клапана, на внутренних стенках средней кишки находятся мелкие складки слизистой оболочки, которые в конце переходят в гладкие стенки задней кишки.
Задняя кишка (rectum) — прямой задний отдел кишечника — оканчивается анальным отверстием. У осетровых рыб нет слепых отростков на границе средней и задней кишки, свойственных другим позвоночным.
Анальное отверстие (anus) — переднее из четырех отверстий анальной зоны, расположенное непосредственно позади пояса брюшных плавников.
Брыжейки. Кишечный тракт в брюшной полости и окружающие его органы соединены со стенками тела и друг с другом мезентериями. Пищевод до места отхождения плавательного пузыря соединен с позвоночником дорзальным мезентерием. Кардиальный и пилорический отделы желудка спаяны брыжейками с долями печени. Селезенка прикреплена брыжейками к кардиальному отделу желудка и далее к двенадцатиперстной кишке. От добавочной доли селезенки и двенадцатиперстной кишки идут мезентериальные спайки к печени, вдоль которых располагаются дольки поджелудочной железы. Спиральная кишка и прямая кишка имеют дорзальный мезентерий, связывающий их сначала с плавательным пузырем, а позади него — с позвоночником. Наконец, гонады и выводные каналы мочеполовой системы своими брыжейками (mesovarium. mesotestis) прикреплены к дорзальной стенке полости тела.
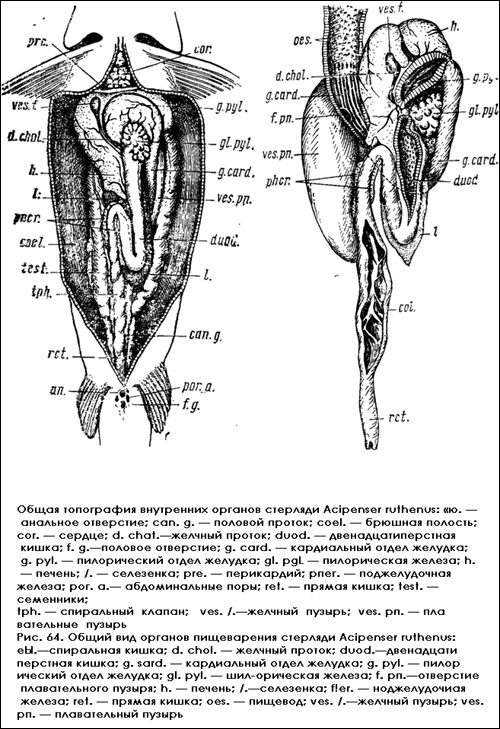
Общая топография внутренних органов стерляди
Мочеполовая система. Строение мочеполовой системы осетровых рыб занимает промежуточное положение между хрящевыми и лучеперыми рыбами. Для всех осетровых рыб характерно наружное оплодотворение и откладка большого количества икры, а также отсутствие клоаки. Оба эти признака связывают осетровых с костистыми и другими лучеперыми рыбами. С другой стороны, у них имеются яйцеводы, открывающиеся воронками в полость тела, что объединяет осетровых с хрящевыми, двоякодышащими рыбами и наземными позвоночными.
Яичники (ovarium) — громадные парные гонады самки — занимают боковые части полости тела почти от самого перикардия до заднего конца целома. Они прикреплены к дорзальной стенке тела брыжейками на всем протяжении справа и слеза от плавательного пузыря. Яичник имеет зернистое строение, он состоит из яйцеклеток одинаковой величины и одинаковой степени развития, так как икрометание происходит одновременно.
Яйцеводы (oviductus) — особые мезонефрические выводные каналы — лежат на наружной стороне гонад в виде широких трубок, открывающихся как у самок, так и у самцов в полость тела широкими воронками на уровне нижней половины гонады. Они соответствуют яйцеводам — мюллеровым каналам — хрящевых и двоякодышащих рыб, но имеют особое происхождение от первичного почечного выводного канала, к которому присоединяется мезонефрическая воронка. Правый и левый яйцеводы на заднем конце сливаются друг с другом и открываются общим отверстием позади анального отверстия.
Семенники (testis) — парные половые железы самцов — занимают, как и яичники, боковые отделы полости тела. Они имеют дольчатое строение и сверху, при рассмотрении вскрытой рыбы, прикрыты большими скоплениями жировых тел. Как и у самок, на наружной стороне железы располагаются воронки яйцеводов, соединяющиеся с широким тонкостенным половым протоком с правой и левой сторон. Внизу парные половые протоки сливаются вместе и открываются половым отверстием, лежащим непосредственно позади анального отверстия.
Мочеточниками (ureter) и семяпроводами (vas deferens) служат первичные почечные протоки. Они начинаются у переднего края почки с наружной стороны отдельными почечными канальцами и образуют общий проток, к которому на уровне заднего конца плавательного пузыря присоединяется воронка яйцевода, образованная у осетровых рыб не мюллеровыми каналами, а мезонефрическим каналом. Через эту воронку и выводной канал выводится наружу жидкость.
При вскрытии общего мочеполового канала видно, как в нем открываются отверстия многочисленных нефридиальных мочевых канальцев.
Семявыносящие канальцы (vas efferens). От семенника, лежащего кнаружи от полового протока, в брыжейках, на которых подвешен семенник, при наблюдении под бинокулярной лупой видны тоненькие, прозрачные выносящие нефридиальные трубочки, идущие вглубь, в почку. Они служат семявыносящими канальцами. Впереди воронки по направлению к верхней доле семенника можно видеть целый букет прозрачных семевыносящих канальцев. Они приносят семя из семенника и общим пучком впадают на уровне воронки в общий семяпровод. Почечные нефридиальные канальцы тонкими трубочками пронизывают почку и впадают в общий проток выводного канала.
Почки располагаются в заднем отделе туловища на дорзальной стенке брюшной полости тела и прикрыты выстилкой брюшины. Для того чтобы их отпрепарировать, необходимо удалить яичники или семенники, подрезав спереди назад брыжейку, прикрепляющую их к стенкам тела. Затем промывают струей воды полость тела и постепенно отчленяют яйцеводы и перитонеальную выстилку полости тела. Отпрепарируют также плавательный пузырь и отодвигают его в сторону. Далее вскрывают ножницами задний отдел брюшной полости рядом с анальным и половым отверстиями и отпрепарируют задние отделы целома, отодвигая в стороны мускулатуру стенок тела.
Почки имеют форму парных лентовидных железистых органов, слившихся друг с другом позади плавательного пузыря по средней линии тела. Они пронизаны кровеносными сосудами, образующими систему почек. Дойдя до заднего конца плавательного пузыря, точки раздваиваются и простираются вперед, оплетаемые разветвлениями задних кардиальных вен, лежащих внутри от почек. Передний конец лентовидных почек доходит до уровня петли желудка на месте перехода кардиального отдела в пилорический.








