Ротоглоточная полость. Для того чтобы вскрыть ротоглоточную полость и рассмотреть передний отдел пищеварительной системы и жаберный аппарат, необходимо отчленить жаберную крышку в месте ее прикрепления к голове, по краю костного панциря, покрывающего голову сверлу, предварительно ознакомившись с ее строением.
Жаберная крышка (operculum) называется оперкулярной крышкой, так как она закрывает все жабры. Изучают ее строение. Это — производная подъязычной жаберной перегородки, лежащей между спиракулярной и первой жаберной щелями. Если сравнить ее с другими жаберными дугами, то становится ясным, что жаберная крышка не что иное, как разросшаяся жаберная перегородка подъязычной дуги. На внутренней стенке жаберной крышки прикреплены жаберные лепестки оперкулярной жабры. Жаберная крышка поддерживается покровными костями и кожными костными чешуями, задний ее край имеет кожистую перепонку, которая служит клапаном всасывающего насоса в дыхательном акте. Жаберные перепонки на вентральной стороне головы с двух сторон прирастают к межжаберному промежутку — истмусу, у белуг они срастаются друг с другом и образуют общую складку. Позади жаберной крышки помещаются наружные жаберные отверстия, окаймленные сзади передним краем пояса конечностей грудных плавников.
Вставляют лезвие ножниц в жаберное отверстие около костного панциря и проводят разрез сзади наперед. На середине расстояния до глаза можно увидеть брызгальце (spiraculum). Проводят разрез через брызгальце дальше вперед и вниз по краю окостенений глазного кольца и вниз на брюшную сторону головы. Затем удаляют с обеих сторон костный и хрящевой пояса грудных плавников. Взяв грудной плавник левой рукой и оттянув его, отчленяют от головы, отрезают мускулатуру грудного пояса, отделяют его от стенки брюшной полости и затем твердыми ножницами перерезают окостенения плечевого пояса посредине. После этого осторожно отделяют вентральные отделы плечевого пояса вместе с плавниками. Далее, вставляют лезвие ножниц в ротовое отверстие и проводят разрез справа и слева так, чтобы он проходил между дорзальным и вентральным отделами жаберных дуг. Оттягивая отрезанные дуги, постепенно отчленяют вентральную стенку жаберного аппарата от его дорзальной стенки по линии разреза. После этого рассматривают сначала ротоглоточную полость, а затем и строение жаберного аппарата.
Ротовое отверстие (stoma) помещается на вентральной стороне головы и имеет форму овального отверстия, расположенного в поперечном направлении; оно окаймлено сверху и снизу мясистыми губами, поддерживаемыми покровными окостенениями.
Ротоглоточная полость (cavum oropharyngeus) резкой границей разделена в поперечном направлении на две части: переднюю ротовую полость — и заднюю — жаберную полость. Первая образует выдвижной челюстной аппарат. Взявшись рукой за верхнюю челюсть, можно вытянуть ротовую воронку из кожного мешка, причем кожная складка вытягивается на небной пластинке.
Выдвижной челюстной аппарат — своеобразное приспособление для схватывания добычи на дне. Он может перемещаться как салазки на большой покровной кости парасфеноида (parasphenoideum), подстилающего крышку ротоглоточной полости. Вытягивающаяся часть челюстного аппарата состоит из небно-квадратного хряща и продолжается далее назад в виде небной пластинки. При оттягивании жаберного аппарата хорошо видна структура небных валиков, располагающихся в поперечном направлении в количестве 7-8 полосок на небной пластинке в 2-3 ряда — на небно-квадратном хряще.
Жаберная полость и жабры. Отворачивают за нижнюю челюсть вентральную стенку ротоглоточной полости и рассматривают строение жаберной полости и жабр. Жаберная крышка костных рыб образована жаберной перегородкой подъязычной дуги, дно глоточной полости — непарными хрящевыми элементами жаберного аппарата, боковые стенки пронизаны справа и слева четырьмя жаберными щелями, отделенными друг от друга жаберными дугами. Пользуясь лупой, рассматривают более подробно строение отдельной жаберной дуги.
Жаберная дуга. Вырезают первую жаберную дугу и рассматривают ее строение более подробно. Верхние концы жаберных дуг прикреплены к небу глоточно-жаберными элементами, а нижние срастаются с подъязычным аппаратом. На внутреннем крае каждой жаберной дуги в два ряда сидят жаберные тычинки, входящие друг в друга; как зубцы шестерни. Один ряд сидит на переднем крае, другой — на заднем. Каждая жаберная тычинка имеет форму конического треугольника. Тычинки выполняют функцию фильтра для удержания пищи. На наружном крае жаберной дуги прикреплены жаберные лепестки, расположенные также в два ряда и отделенные друг от друга жаберными перегородками.
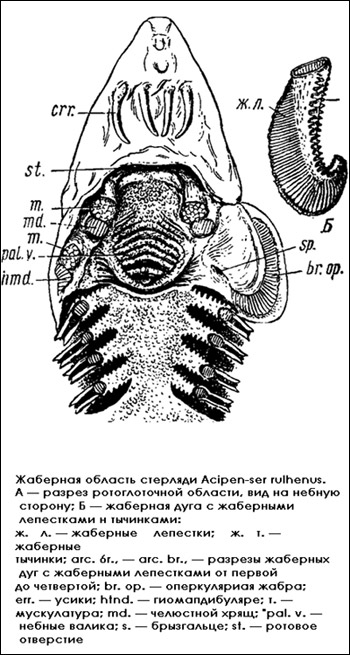
Жаберная область стерляди
Жаберные перегородки не достигают края жаберных лепестков (в отличие от акуловых рыб, где жаберные перегородки простираются до краев жаберных отверстий). Каждый жаберный лепесток имеет форму листовидной перегородки, в которой помещается петля жаберных капилляров, составленная приносящим и выносящим жаберными сосудами. Жаберная дуга с двумя рядами жаберных лепестков на наружной и внутренней сторонах образует одну целую жабру рыб (holobranchia).
Передний ряд жаберных лепестков располагается на внутренней поверхности жаберной крышки, является полужаброй подъязычной дуги и получает название оперкулярной жабры (branehia opercularis); она ограничивает переднюю стенку первой жаберной щели. На задней стенке передней жаберной щели лежит второй ряд жаберных лепестков, который спаян жаберной перегородкой первой жаберной дуги со следующим рядом жаберных лепестков, являющихся передней стенкой второй жаберной щели. Второй ряд жаберных лепестков вместе с третьим рядом составляет первую жабру. Вторая, третья и четвертая жабры также состоят из двух рядов лепестков каждая. Последняя, пятая, жаберная дуга не несет жаберных лепестков, вплотную прилегает к плечевому поясу и скрыта под кожей, поэтому задняя стенка пятой жаберной щели ограничена передним краем плечевого пояса.
Акт дыхания у осетровых рыб, как и у других костных рыб, носит название оперкулярного дыхания. При оттягивании жаберной крышки в стороны в околожаберной полости между жаберной крышкой и жабрами образуется пространство с пониженным давлением, куда всасывается вода из ротоглоточной полости через жаберные отверстия. В ротовую же полость вода всасывается через рот при растяжении всего висцерального аппарата. При прижимании жаберной крышки вода выталкивается из околожаберной полости через наружные жаберные отверстия.
Кровеносная система. Кровеносная система осетровых рыб обладает типичными чертами кровообращения рыб. Сердце подразделено на четыре отдела (артериальный конус, желудочек, предсердие и венозный синус) и содержит венозную кровь, которая по брюшной аорте и приносящим жаберным сосудам приносит кровь к жабрам, где происходит ее окисление. Артериальная кровь собирается в выносящие жаберные сосуды и вливается в спинную аорту, снабжающую своими ветвями все органы. Основными венозными стволами, приносящими кровь к сердцу, являются передние и задние кардинальные вены. Пищеварительная система имеет обособленную венозную систему воротную вену печени, приносящую кровь к печени, где происходит фильтрация крови через систему капилляров.
Изучение кровеносной системы надо начинать с сердца, для чего следует отпрепарировать околосердечную сумку, располагающуюся впереди брюшной полости тела, между грудными плавниками.
Перикардиальная полость, или околосердечная сумка — обособившаяся передняя часть брюшной полости тела. Она отделена от нее соединительнотканной перегородкой, лежащей в поперечном направлении впереди печени. Перикардиальная полость имеет форму треугольника, направленного вершиной вперед, что можно было видеть при общем вскрытии. В этой треугольной полости располагается сердце рыб. Полость перикардия выстлана брюшиной (перитонеумом), состоящей из двух оболочек: наружная называется перикардием, а внутренняя, прилегающая к сердцу и прикрепляющая его к стенке тела,- эпикардием.
Сердце, как указано выше, состоит из четырех отделов, обладающих способностью пульсации. Сначала рассматривают отделы сердца, а потом разбираются в его внутреннем строении на разрезе. Передним отделом является артериальный конус (conus arteriosus) в виде короткой трубки, лежащей впереди мускулистого желудочка. Желудочек (ventriculus) имеет трапециевидную форму с плоским и даже вогнутым основанием, направленным вниз, а вершиной — вперед. Так же, как и артериальный конус, он имеет толстые мускулистые стенки. Наружная поверхность желудочка и артериального конуса у всех осетровых рыб покрыта почти сплошь пузыревидными расширениями железистого характера, это типичная для осетровых лимфоидная железа, представленная отдельными дольками. Это можно видеть на фронтальном разрезе сердца. В глубине, под желудочком, располагается задний отдел сердца рыб — предсердие (atrium), имеющий вид сморщенного мешка с мускулистыми стенками из густопереплетенных волокон. Совсем в глубине околосердечной сумки предсердие общим широким отверстием сообщается с самым задним отделом сердца — венозным синусом (sinus venosus), принимающим венозную кровь со всего тела рыбы. Если сделать фронтальный разрез желудочка сердца, начиная от вершины, направленной вниз, и далее вперед через артериальный конус, станут видны толстые мускулистые стенки желудочка с радиальным расположением переплетающихся мышечных волокон. Внутренняя полость желудочка узкая, с лакунарными карманами, открывающимися в желудочек. На задней стенке этой полости имеется широкое атриовентрикулярное отверстие (ostium atrioventncularis), открывающееся в предсердие. На границе между полостью желудочка и артериального конуса стоят 3-4 листовидные лопасти, переграждающие обратное возвращение крови в желудочек. В самом артериальном конусе расположены три ряда полулунных клапанов (valvula semilunaris): два ряда внизу, а третий ряд выше — у основания брюшной аорты.








